Life chirality problem can be solved in more steps. A most general question is, why the organisms are accepting the matter and energy in different ways, at second we should explain, why the energy and building blocks molecules differs in chirality, while the third (apparently more difficult question) is, why just the D-sugars and L-amino-acids are accepted by living creatures, but not vice-versa.
The first question isn't so apparent in this moment, but it's connected to dual character of stable forms of matter, which are divided into fermions (female) and boson (male) particles (elements). Both males, both females are occupying their homes/shells/nests, which can be interpreted as sinuses formed by building blocks. In social species, the females are collecting a building blocks, while males are foraging another energy sources preferably. The second question is related to problem, why the building blocks are usually collected from local sources or proximity, while food and energy sources are collected from longer distance and the third problem is related to question, why females tends to forage a food and building blocks, while the males are specialized in collecting of energy sources preferably, usually from outside of their homes.
We can observe an apparent chirality in SU(3) Young polyominos defined in 3D by 2x2x2 Jourdan triade matrix of mutual male/female, mass/energy and proximity/distance dualities, which follows from the fact, the males are movable like energy supplies (a prey) and therefore they appear at the longer distance from homes. This chirality is repeated in many fractal levels, because the fermion particles are serving as a food/energy supplies for subsequent nested levels of food chain.
By AWT the concept of CP invariance violation and chirality of life are closely related by concept of foam, composed of walls/membranes of nonzero thickness. The particles are formed by standing wave packets, spreading along surfaces of these membranes, but the situation of inner and outer surface isn't always very same.


The particles formed by inner gradients of Aether foam membranes are always of higher space-time curvature and mass/energy density, then these outer ones. During condensation of foam the particles formed by inner surfaces will condense into fermions - while the rest of matter will evaporate instead into particles of energy, i.e. the bosons. While total chirality is retained, the evaporating bosons are formed by super-symmetric anti-particles preferably. By AWT during Universe formation (a Big-Bang event) a symmetric graviton foam was formed first. This foam was formed by membranes with (nearly) no preference in concave or convex curvature. The similar foam is formed at the beginning of supercritical fluid condensation.


Whenever the density of graviton foam has increased, a second phase transition has occurred, so called the inflation. During this a system was formed, consisting of nested density fluctuations separated by membranes, composed by adjacent pair of surface gradients. After then the condensation of matter has separated into two branches. A density fluctuations formed by inner walls of foam branes have continued in condensation, forming a dense particles of observable matter. While the second portion of particles has evaporated instead into anti-symmetric bosons, which formed the sparse streaks of dark matter.
We can assume, the very same process has repeated later during life formation. By AWT the life was formed in so-called liposomes, formed by fragmentation of tiny oily droplets at the stormy surface of ancient oceans. Every liposome is very small (i.e. of high surface curvature) and it has a double surface, formed by hydrophobic membranes. In such system, the surface tension phenomena cannot be neglected, because the highly concave surface exhibits so-called superhydrophobicity, so it repels the carbohydrate molecules with many -OH functional groups, which are collected preferably inside of liposome. While the building blocks of organic matter, i.e. the amino-acids with hydrophobic radicals should adsorb at the outer surface of liposome, preferably. This forms natural condition for spatial arrangement of t-RNA components at the phase interface of liposomes. They should probably form spontaneously in thin dispersion of lipids in solution of aminoacids and ribose mixture.
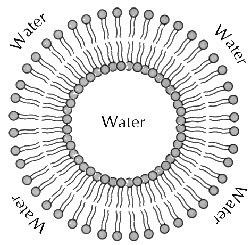
The chirality of liposome follows from so called Hairy ball theorem: every surface of droplet should contain an at least two quantum vortices (an analogy of Cooper pairs condensation in solid state physics) and these vortices are of opposite chirality due the geometry reasons. So that the chiral molecules should separate on both surfaces of liposome by its helicity. For liposomes of larger diameter there is higher number of quantum vortices just at the outer surface of liposome, which is always slightly larger due the nonzero thickness of lipid bilayer (a liposome membrane).
By such way, a chiral organic molecules should obtain their distribution from the very beginning of liposome formation, i.e. it's not accidental, one-shot process depending on panspermia event - but in quite reversible process, which should be replicated in experiments both in supercritical fluid or boson condensate systems, both in liposome dispersions in laboratory conditions. This explanation supports the contemporary models of terrestrial life formation, by which sponges are one of the first multicellular organisms. By AWT the life was formed from liposome foam and the ancient sponges are just another nested phase of it (i.e. sponge of cellular sponge by the same way, like atom nuclei are formed by quantum foam of smaller strings, etc..).
Energy spreads by slowest speed just along foam surfaces, so AWT prefers atemporal fractal foam structures during gradual evolution of complexity over other geometries and the above findings are consistent with this concept.


9 comments:
By AWT the chiral compounds by the inner and outer surface of liposome membranes during formation of life is a direct analogy to separation of chiral portions of matter during formation of Universe.
http://aetherwavetheory.blogspot.com/2009/03/is-dark-matter-composed-of-antimatter.html
Third law of prediction: "Any sufficiently advanced technology is indistinguishable from magic."
Arthur C. Clarke: Profiles of the Future, 1973.
How AWT could explain prion disease?
I presume prions are the chiral version of a healthy protein called PrP 33-35. Whereas healthy PrP is normally located on the cell's SURFACE, the mirror D-protein goes astray and end up in the cytosol, the liquid found INSIDE cells (basically like common D-sugars). In addition, chiral proteins are sticky because of their opposite dipole charge (..from basically the same reason, like the electrons are attracted to positrons...) and they adhere to almost every normal L-protein inside of cells
So when PrP misfolds, the results are disastrous. Because D-proteins aren't metabolized by common enzymes, they tend to cumulate in food chain of living organisms. This is why we shouldn't eat dogs, cats and other predators (including some fishes, like merlin, shark or tuna), not saying about cannibalism. The cows are especially sensitive to cumulation of prions, because as a herbivores they've no enzymes for dissolution of animal proteins at all.
The pigment array in thylakoid lamellas i.e. quantasomes appear pretty similar to quantum dots arrays. Each quantasome contains about 230 to 300 chlorophyll molecules. They're regularly spaced in 150 x 180 A lattice, like quantum vortices in superconductors. All the molecules in each of these photo-synthetic units are spaced and oriented in such a way, captured photons are transferred from molecule to molecule by inductive resonance and the energy absorbed is transferred to as exciton.
Because the quantum boost photosynthesis is relatively small (~15%), experiments have demonstrated, that the presence of the quantasome particles in chloroplast membrane is not a necessary condition for photoreduction activity of chloroplasts [J. Mol. Biol., 27, 323 (1967)]. In more primitive prokaryotes cells pigments are distributed uniformly on or in the thylakoid lamellae.
Homochirality results from boiling solutions containing a racemic mixture of enantiomorphous
crystals due to dissolution-crystallisation induced by a temperature gradient
Life began with a planetary mega-organism
We could illustrate the above explanation with following analogy: the molecules are behaving like an insect with four legs arranged into vortices of tetrahedron. One of its leg is oiled and as such hydrophobic and the another ones are hydrophilic. To give a chirality to such a molecule, we can imagine, the whole body of insect is hydrophilic (sugar molecule) or hydrophobic (aminoacids molecules). The hydrophilic molecules would climb more easily around positively curved hydrophilic surface represented with sphere (biological cell formed with lipidic bilayer), where the higher number of hydrophilic parts of its body would get into contact with the surface of sphere. Whereas the hydrophobic molecules would prefer the internal negatively curved surface of the sphere from the same reason. For hydrophilic molecules it would be advantageous to have oiled leg oriented outside of sphere, for second ones toward center of sphere. Their separation would increase with decreasing radius of the cell.
Probably.
Chiral recognition and selection during the self-assembly process of protein-mimic macroanions
[Development of Chiral and Achiral Supercritical Fluid Chromatographic methods for the characterisation of ophthalmic drug substances and drug products](http://doras.dcu.ie/21398/1/Adrian_Marley_Final_Thesis_07_Sept_2016__AMS.pdf)
Post a Comment